ISSN: 1839-9940

 Global reach, higher impact
Global reach, higher impactJ Genomics 2025; 13:6-9. doi:10.7150/jgen.103481 This volume Cite
Short Research Paper
Draft genomes and assemblies of the ectomycorrhizal basidiomycetes Scleroderma citrinum hr and S. yunnanense jo associated with chestnut trees
Haolin Zhang1, Jiayi Han1, Richard D. Hayes2, Kurt LaButti2, Igor Shabalov2, Anna Lipzen2, Kerrie Barry2, Igor V. Grigoriev2,3, Qing Zhang1, Qingqin Cao1, Huchen Li1 ![]() , Francis M. Martin4,5
, Francis M. Martin4,5 ![]()
1. Beijing Key Laboratory of New Techniques in Agricultural Application, Beijing University of Agriculture, Beijing 102206, China.
2. U.S. Department of Energy Joint Genome Institute, Lawrence Berkeley National Laboratory, Berkeley, CA 94720, USA.
3. Department of Plant and Microbial Biology, University of California Berkeley, Berkeley, CA 94720, USA.
4. Université de Lorraine, INRAE, UMR 1136 Interactions Arbres/Microorganismes, 54280, Champenoux, France.
5. Northwest Institute of Eco-Environment and Resources, Lanzhou, China.
Received 2024-9-10; Accepted 2024-11-20; Published 2025-1-1
Abstract

The earthball Scleroderma, an ectomycorrhizal basidiomycete belonging to the Sclerodermataceae family, serves as a significant mutualistic tree symbiont globally. Originally, two genetically sequenced strains of this genus were obtained from fruiting bodies collected under chestnut trees (Castanea mollissima). These strains were utilized to establish in vitro ectomycorrhizal roots of chestnut seedlings. The genome sequences of these strains share characteristics with those of other ectomycorrhizal species in Boletales order, including a restricted set of genes encoding carbohydrate-active enzymes. The genome sequences presented here will aid in further exploring the factors contributing to the establishment of ectomycorrhizal symbiosis in chestnut trees.
Keywords: Boletales, ectomycorrhizal symbiosis, genome, Scleroderma, Sclerodermataceae.
Scleroderma, commonly known as earthballs, is a widely distributed ectomycorrhizal gasteromycetes genus that produces large, noticeable sporocarps or earthballs in various forest environments and areas adjacent to forests [1]. The ectomycorrhizal status of S. citrinum and S. yunnanense has been confirmed and symbiosis can be established in in vitro experimental systems [2]. Scleroderma citrinum belongs to the family Sclerodermataceae. This mushroom appeared early in the fruiting succession of ectomycorrhizal fungi. It is the primary colonizer of mining waste, enabling it to spread rapidly and colonize young root systems of numerous tree species. The genus Scleroderma is found worldwide, and S. citrinum is commonly recorded in temperate regions of the Northern Hemisphere (Fig. 1A) [3]. Several host genera have been reported for Scleroderma, including poplars and eucalypts [4,5]. Multiple studies have investigated the symbiotic relationships between Scleroderma species and their host plants, focusing on the growth and nutritional benefits of this relationship [3,6-8]. Recently, the S. citrinum mycorrhizosphere has been studied as a model system to examine the impact of ectomycorrhizal symbiosis on the taxonomic and functional diversity of bacterial communities involved in mineral weathering [3,9,10].
Basidiocarps (A) and vegetative mycelial cultures (B) of S. citrinum and S. yunnanense. A, natural habitate of S. citrinum hr. (B) Growth of S. citrinum hr and S. yunnanense jo strains on MMN, P20, and PDA solid media 1 or 30 days after inoculation.

In October 2018, S. citrinum hr. and S. yunnanense jo. strains were isolated from fruiting bodies growing under chestnut trees (Castanea mollissima) in Huairou and Jianou, China, respectively. These strains were cultured on agar plates containing the P20 medium at 25°C, and subsequent mycelial (monosporal) cultures were used to remove contaminants until identical mycelia were obtained. The identity of the purified strains was confirmed by sequencing the rDNA ITS region using the primers ITS4 and ITS5[11]. After molecular characterization, both strains were deposited at Beijing Key Laboratory of New Techniques in Agricultural Application with accession numbers 0322 for S. citrinum hr and 0323 for S. yunnanense jo. The growth of both strains on P20, MMN, and PDA media was compared, revealing that P20 medium was the most optimized for both strains (Fig. 1B).
Fresh mycelia were obtained from fungal colonies grown on the P20 medium and were snap-frozen in liquid nitrogen. The samples were subsequently ground using a TissueLyser LT (QIAGEN). DNA was isolated following the CTAB extraction protocol [12], and total RNA was isolated using an RNAeasy extraction kit (QIAGEN), according to the manufacturer's instructions. During the extraction procedures, RNase A or DNase I (both from Thermo Fisher) were used to purify DNA or RNA, respectively. Approximately 200 μg of DNA and 10 μg of total RNA were extracted from each strain.
The S. citrinum hr and S. yunnanense jo v1.0 genomes were sequenced from 100 μg of genomic DNA using the Pacific Biosciences sequencing platform (>10kb PacBio libraries with Blue Pippin size selection), assembled with Falcon v. 0.0.8[13], polished with Arrow version SMRTLINK v8.0.0.80529 (https://www.pacb.com/support/software-downloads), and annotated using the MycoCosm developed by JGI[14], information of genomic portals can be found in https://mycocosm.jgi.doe.gov/Sclcihr1 and https://mycocosm.jgi.doe.gov/Sclyun1. The Whole Genome Shotgun project has been deposited in GenBank under BioProjects PRJNA711063 and PRJNA1080752. To support gene prediction, mRNA sequences were obtained using Illumina RNA-Seq data assembled using Trinity v2.11.0[15].
The assembly size of the Chinese strain hr of S. citrinum (77,47 Mbp), was higher than that of the previously sequenced European strain S. citrinum Foug A (i.e., 56,14 Mbp), whereas the number of genes was lower (10323 genes versus. 21012 genes) [16]. We identified polymorphic content within the assembly and annotation of strain hr, which reflected significant separation of alleles. Many scaffolds were observed to be highly similar to larger scaffolds and were predicted to constitute alternate or secondary haplotypes. The assembly size of the S. yunnanense genome (45,55 Mbp) was lower than that of S. citrinum genomes, despite the fact that the number of genes was very similar between the two Chinese strains, specifically 10,323 versus. 10,194.
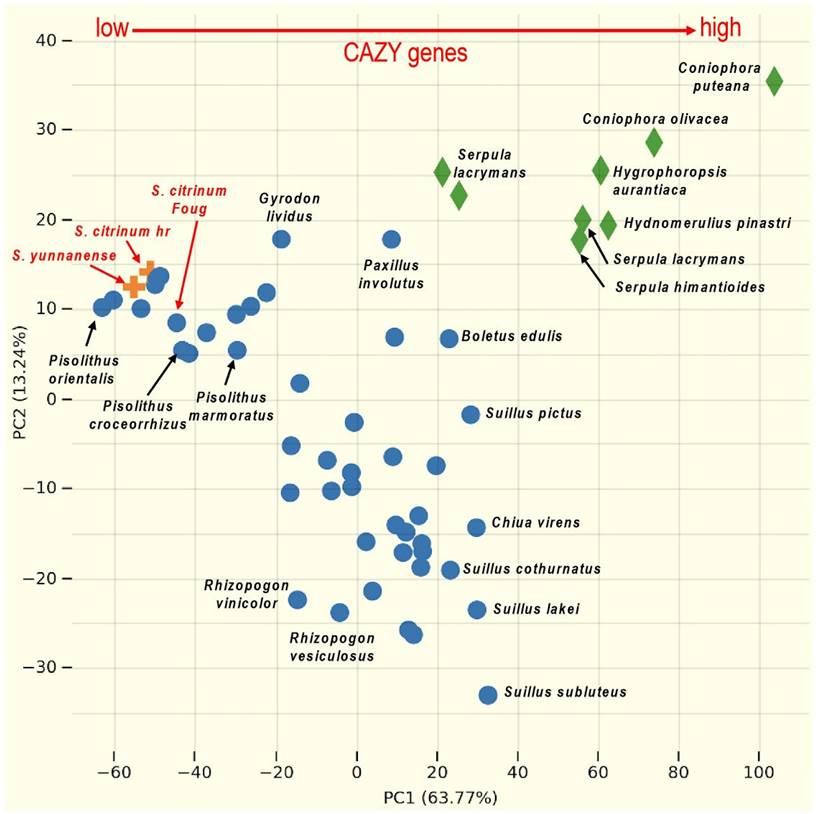
A restricted set of genes encoding carbohydrate-active enzymes (CAZymes) is a major trait in the evolution of ectomycorrhizal fungi from saprotrophic ancestors [16]. The genomes of S. citrinum hr and S. yunnanense contain 244 CAZyme genes, whereas the S. citrinum FougA genome encodes 257 CAZyme genes. This repertoire of CAZyme genes acting on lignocellulose is much lower than that of their saprotrophic relatives in Boletales, such as Coniophora and Serpula species, as well as ectomycorrhizal fungi, such as Suillus and Boletus spp. (Fig. 2). In contrast, their CAZyme repertoire is very similar to that of Pisolithus species, which are also found in sandy soils with low soil organic matter content.
Differential distribution of CAZyme gene copy numbers in the genomes of 92 species of saprotrophic and ectomycorrhizal fungi in Boletales. Principal coordinate analysis of the total CAZyme gene copy numbers identified in saprotrophic and ectomycorrhizal fungi. Each symbol corresponds to the genome of the Boletales species available in the JGI MycoCosm database (mycocosm.jgi.doe.gov). The lifestyle of the sequenced species is indicated by different colored species names (green for saprotrophic species and blue and red for ectomycorrhizal species). Divergent distributions of CAZyme gene sets in various mycorrhizal lifestyles with an increasing repertoire of CAZyme from ectomycorrhizal Scleroderma species (in red) to saprotrophic brown rotters (higher; from left to right). Principal component analysis (PCA) was performed using the MycoCosm PCA tool (mycocosm.jgi.doe.gov). The CAZyme gene repertoires were obtained after semi-manual curation of protein sequences by the CAZy team (www.cazy.org)[17].
The current set of genome sequences will enable additional research into the molecular factors that drive the formation of ectomycorrhizal symbiosis in chestnut trees.
Acknowledgements
This project was part of the JGI CSP '1KFG: Deep sequencing of ecologically relevant Dikarya' (# 10.46936/10.25585/60001060) (PI: FM), conducted by the US DOE Joint Genome Institute, a DOE Office of Science User Facility, supported by the Office of Science of the US DOE under contract no. DE- AC02-05CH11231. This work was also supported by a grant from the French National Research Agency (ANR) as part of the ''Investissements d'Avenir'' program (ANR-11-LABX-0002-01, Lab of Excellence ARBRE) (to FM) and grants from the National Natural Science Foundation of China (32472671 & 32002019) to HL.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Theodorou C, Reddell P. In vitro synthesis of ectomycorrhizas on Casuarinaceae with a range of mycorrhizal fungi. The New phytologist. 1991;118(2):279-288
2. Freire CG. et al. First record of in vitro formation of ectomycorrhizae in Psidium cattleianum Sabine, a native Myrtaceae of the Brazilian Atlantic Forest. PloS one. 2018;13(5):e0196984-e0196984
3. Calvaruso C. Impact of ectomycorrhizosphere on the functional diversity of soil bacterial and fungal communities from a forest stand in Impact of ectomycorrhizosphere on the functional diversity of soil bacterial and fungal communities from a forest stand in relation to nutrient mobilization processes. Microbial Ecology. 2007;54(3):567-77
4. Chen Y.L. et al. Mycorrhizal status of Eucalyptus plantations in south China and implications for management. Mycorrhiza. 2007;17(6):527-535
5. Zhu Q. et al. Rhizosphere bacterial and fungal communities succession patterns related to growth of poplar fine roots. Sci Total Environ. 2021;756:143839
6. Itoo ZA, Reshi ZA. Influence of ectomycorrhizal inoculation on Pinus wallichiana and Cedrus deodara seedlings under nursery conditions. Frontiers in biology. 2014;9(1):82-88
7. Colpaert J.V. et al. Van Assche, Carbon and nitrogen allocation in ectomycorrhizal and non-mycorrhizal Pinus sylvestris L. seedlings. Tree Physiol. 1996;16(9):787-93
8. Sousa N.R. et al. Ectomycorrhizal fungi as an alternative to the use of chemical fertilisers in nursery production of Pinus pinaster. Journal of environmental management. 2012;95:S269-S274
9. Uroz S. et al. Effect of the Mycorrhizosphere on the Genotypic and Metabolic Diversity of the Bacterial Communities Involved in Mineral Weathering in a Forest Soil. Applied and Environmental Microbiology. 2007;73(9):3019-3027
10. Uroz S. et al. Functional profiling and distribution of the forest soil bacterial communities along the soil mycorrhizosphere continuum. Microb Ecol. 2013;66(2):404-15
11. Horton T.R, Bruns T.D. The molecular revolution in ectomycorrhizal ecology: peeking into the black-box. Mol Ecol. 2001;10(8):1855-71
12. Moller E.M. et al. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992;20(22):6115-6
13. Chin C. et al. Phased diploid genome assembly with single-molecule real-time sequencing. Nature methods. 2016;13(12):1050-1054
14. Grigoriev I.V. et al. MycoCosm portal: gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014;42(Database issue):D699-704
15. Friedman N. et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nature biotechnology. 2011;29(7):644-652
16. Kohler A. et al. Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists. Nat Genet. 2015;47(4):410-5
17. Drula E. et al. The carbohydrate-active enzyme database: functions and literature. Nucleic Acids Res. 2022;50(D1):D571-D577
Author contact
![]() Corresponding author: Huchen Li; huchen.liedu.cn and Francis M. Martin, francis.martinfr.
Corresponding author: Huchen Li; huchen.liedu.cn and Francis M. Martin, francis.martinfr.