ISSN: 1839-9940

 Global reach, higher impact
Global reach, higher impactJ Genomics 2018; 6:1-8. doi:10.7150/jgen.23248 This volume Cite
Research Paper
Genetic variation in N- and C-terminal regions of bovine DNAJA1 heat shock protein gene in African, Asian and American cattle
Oyeyemi O. Ajayi1, Sunday O. Peters2,3, Marcos De Donato1,4, F. Denis Mujibi5, Waqas A. Khan1,6, Tanveer Hussain1,7, Masroor E. Babar8, Ikhide G. Imumorin1,9,10 ![]() , Bolaji N. Thomas11,
, Bolaji N. Thomas11, ![]()
1. Animal Genetics and Genomics Laboratory, International Programs, College of Agriculture and Life Sciences, Cornell University, Ithaca, NY 14853
2. Department of Animal Science, Berry College, Mount Berry, GA 30149
3. Department of Animal and Dairy Science, University of Georgia, Athens, GA 30602
4. Departamento Regional de Bioingenierias, Instituto Tecnologico y de Estudios Superiores de Monterrey, Queretaro, Mexico
5. Usomi Ltd., PO Box 105086-00101, Ushirika Road, Karen, Nairobi, Kenya
6. Department of Biotechnology, University of Sargodha, Sargodha, Pakistan
7. Department of Molecular Biology, Virtual University of Pakistan, Lahore, Pakistan
8. African Institute for Biosciences Research and Training, Ibadan, Nigeria
9. School of Biological Sciences, Georgia Institute of Technology, Atlanta, GA 30332
10. African Institute for Biosciences Research and Training, Ibadan, Nigeria
11. Department of Biomedical Sciences, Rochester Institute of Technology, Rochester NY, 14623
Received 2017-10-9; Accepted 2017-11-18; Published 2018-1-1
Abstract

DNAJA1 or heat shock protein 40 (Hsp40) is associated with heat adaptation in various organisms. We amplified and sequenced a total of 1,142 bp of bovine Hsp40 gene representing the critical N-terminal (NTR) and C-terminal (CTR) regions in representative samples of African, Asian and American cattle breeds. Eleven and 9 different haplotypes were observed in the NTR in Asian and African breeds respectively while in American Brangus, only two mutations were observed resulting in two haplotypes. The CTR appears to be highly conserved between cattle and yak. In-silico functional analysis with PANTHER predicted putative deleterious functional impact of c.161 T>A; p. V54Q while alignment of bovine and human NTR-J domains revealed that p.Q19H, p.E20Q and p. E21X mutations occurred in helix 2 and p.V54Q missense mutation occurred in helix 3 respectively. The 124 bp insertion found in the yak DNAJA1 ortholog may have significant functional relevance warranting further investigation. Our results suggest that these genetic differences may be concomitant with population genetic history and possible functional consequences for climate adaptation in bovidae.
Keywords: Cattle, Hsp40 gene, climate adaptation, genetic diversity
Introduction
Heat shock proteins of the 40-kDa type (Hsp40) provide protection from proteotoxic stress by repairing stress-damaged proteins and suppressing protein aggregation [1-4], implying that this disruption of proteostasis has both functional and clinical relevance [5,6]. DnaJ proteins are co-chaperones of the DnaK/Hsp70, with whom they carry out their co-chaperone function in facilitating the correct folding of client proteins [7,8]. Published reports have suggested that DnaJs are the primary mediators of this binding to client proteins, presenting them to DnaK/Hsp70 afterwards [9,10], although some exhibit self-chaperone activity [11]. The extensive diversity of architectural domains and subcellular localizations in DnaJs reveals the presence of a J-domain signature that is important for stimulating ATPase activity [12] and for binding DnaK/Hsp70. Details of the functionality of specific J protein remains elusive, even though sequence analysis has highlighted the diversity of the J protein family [5].
The Hsp40 family is a large group of proteins, classified on the basis of molecular weight (~ 40 kDa), with structural and functional diversity [2], yielding 3 subtypes (types I, II, and III), whose sub-classifications are based on the degree of domain conservation, when compared to those of Escherichia coli DnaJ [13]. The J domain at the N-terminus of type I is a disordered glycine-phenylalanine (G/F)-rich region, possibly serving as a linker between the J-domain and the C-terminal region of DnaJ proteins, 2 zinc-finger-like motifs, and a conserved carboxyl-terminal domain (CTD) [14]. Type II Hsp40s on the other hand, lack the zinc-finger-like motifs while only conserved J domains are found in type III Hsp40s. Interactions between Hsp40 and Hsp40-like proteins with their co-chaperone (Hsp70s) are believed to be mediated by J domain as the major binding site [15]. The J domain architecture includes 4 α-helices (helices I-IV), with a loop region containing a highly conserved tripeptide of histidine, proline, and aspartic acid (HPD motif) located between helices II and III [16]. Studies of binding inhibition using peptides suggest that the J domain in yeast Hsp40 called Ydj1 has minimal Hsp70-binding site between amino acids 2 and 35, which includes helices I and II, and the HPD motif [17], allowing the HPD motif to help alter the orientation of the charged residues in helix II to produce correct interaction of helix II with the ATPase domain of a partner Hsp70 [18,19]. Additionally, type I and II Hsp40 protein contain a C-terminal peptide binding fragment which recognizes and bind hydrophobic side chains of denatured polypeptides [20], transferring non-native polypeptides to the Hsp70 peptide-binding domain in the process.
The critical roles of the N and the C-terminus region of the DnaJ proteins in chaperone function, alongside their partner Hsp70s suggest that mutations in these regions could directly or indirectly abrogate chaperoning functions, with attendant consequences for disease-associated protein misfolding and unmitigated cellular stress. To test this hypothesis, we sequenced the N- and C-terminal regions of the bovine DNAJA1 heat shock protein gene in 15 cattle breeds from Asia, Africa and North America and compared to yak to better understand genetic diversity in the bovine DNAJA1 gene in Bovidae. Our goal is to elucidate the underlying molecular footprints in the DNAJA1 gene that may be characteristic of evolutionary and functional differences in geographically separated breeds of cattle.
Materials and Methods
Animal sampling and DNA isolation
Total of 86 blood samples from 15 geographically separated cattle breeds and yak were collected from Africa (Nigeria), Asia (Pakistan) and United States of America. They were made up of Pakistani breeds: Achai (n = 5), Cholistani (n = 5), Bhagnari (n = 5), Dajal (n = 7), Dhanni (n = 7), Lohani (n = 5), Narimaster (n = 5), Red Sindhi (n = 5), Sahiwal (n = 5), Therporter (n = 5) and yak (n = 4); American breed: Brangus (n = 7); Nigerian breeds: Muturu (n = 6), N'dama (n = 5), Sokoto Gudali (n = 5), and White Fulani (n = 4). About 3 ml of blood collected from each animal was processed for genomic DNA isolation, yield and quality, as previously described [21].
PCR amplification and SNP discovery
Two primer pairs (F: 5'-GTTGAAAGTCGGTTAAAATTGAGC-3' and R: 5'-TCATGAAGCCGTGTATACTCCC-3') and (F: 5'-GGAAGAAAGGGCATCTTTTGAAAT-3' and R: 5'-TTGGCACTTCTGGCAATTCAG-3') were designed using NM_001015637.1 as reference sequence to amplify 619 bp spanning exon 2, introns 2 and 3 in the N-terminal domain and 523 bp spanning exon 7, intron 7 and exon 8 in the C-terminal region of the cattle DNAJA1 gene respectively. PCR amplifications were carried out in a total reaction volume of 20µl containing approximately 50ng DNA, as previously described (Adefenwa et al. 2013) and the following cycling conditions: denaturation at 95oC for 4 min, followed by 35 cycles of denaturation at 94oC for 30 s, annealing at 53oC (for N-terminal region) and 54oC (for C-terminal region) for 30 s, extension at 72oC for 1 min, and extended elongation at 72oC for 10 min.
Agarose gel electrophoresis was used to detect amplified products on a 1.5% ethidium bromide-stained gel, and band sizes scored with a GENEMate Quanti-Marker 100 bp DNA ladder (BioExpress, Kaysville, UT, USA). Amplified PCR amplicons were purified with the Zymo DNA Clean and ConcentratorTM kit (Zymo Research Corp., Irvine, CA, USA) and sequenced at the Cornell University Genomics Core Facility.
SNP identification and genotyping
Sequence bases were called and assembled, as previously described [21], while single nucleotide polymorphisms (SNPs) were identified with Codoncode Aligner 3.7.1 software (http://www.codoncode.com/aligner) and DnaSP version 5.10.01 [22]. Using NCBI accession number NM_001015637.1 as reference, we detected SNPs, as described previously [21], utilizing the nomenclature for sequence variation (http://www.hgvs.org/mutnomen). Genotypes were obtained from analyzing and comparing electropherograms.
Sequence analysis
To calculate index of sequence variation, haplotype structure, including numbers of polymorphic sites and haplotypes, DnaSP software version 5.10.01 [22], while PANTHER [23] was used to to assess in-silico functional analysis of missense mutations while alignment of the nucleotide sequences was done using ClustalX [24].
Results
Two primer pairs were designed to amplify 619 bp in the N-terminal region spanning exon 2, introns 2 and 3 and of 523 and 717 base pairs in the C-terminal regions of domestic cattle and yak respectively) spanning exon 7, intron 7 and exon 8 of the bovine DNAJA1 gene (Figure 1). BlastN analysis predicted 99% similarity between the N-terminal region and the reference sequence (NM_001015637.1). Sequence alignment of the N-terminal region using ClustalW identified 5 polymorphic sites in Asian and African breeds and two polymorphic loci in American breed (Brangus). The two polymorphic sites identified in Brangus were also identified in both African and Asian breeds. Interestingly, all the SNPs identified in this study occurred in exons 2 and 3, followed by the presence of invariant sites in intron 2.
Upper band corresponds to 717 bp region observed in Yak; lower band corresponds to 523 bp region of the C-terminal region of domestic cattle, corresponding to peptide binding domain in cattle species.
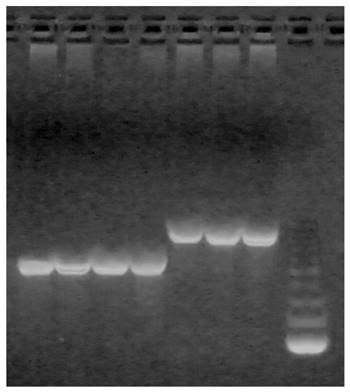
The C-terminal region appears to be largely conserved among African, Asian and American breeds of cattle as well as in yak. However, there were a few notable differences between the consensus cattle and yak sequences with respect to the C-terminal region (Figure 2). Alignment of the consensus sequences of yak and domestic cattle identified the following SNPs: c. 914A>G in exon 7; c. 1035A>T and c.1038T>C in exon 8 of yak DNAJA1 gene using NM_001015637.1 as reference (Figure 3). Although c.914A>G synonymous mutation resulted into p. C271 silent mutation, c.1035 A>T and c. 1038 T>C non-synonymous mutation resulted in p. I310F (amino acid change from isoleucine to phenylalanine at position 310) and p. Y311H (amino acid change from tyrosine to histidine at position 311) missense mutation respectively. Notably, a 124 bp insertion in intron 8 was observed in yak but absent in the cattle DNAJA1 gene, and was also not found in the bovine genome database (Figure 2).
Carboxylic terminal region in bovine and yak. Red shows polymorphic region between bovine and yak DNAJA1 gene locus. Yellow region is present in the bovine genome but absent from amplified cattle sequence aligned to yak. Green is 124 base pair insertion observed in yak DNAJA1 gene but completely absent in cattle. Regions containing arrows are those with nucleotide changes in the coding regions (first arrow on A->G SNP in yak occurred in exon 7 while the second and third arrows on A->T and T->C respectively occurred in exon 8). Red without arrows shows nucleotide changes in intron 7. Stars with arrows mark the beginning and ending of exon 7 and exon 8 respectively. Level of similarity between cattle and yak for DNAJA1 is 98%. Alignment was carried out using ClustalX

SNP positions on the aligned DNAJA1 N- terminal J domain region in cattle aligned with human (NP_001015637.1), mouse (NP_001530.1), dog (NP_001158143.1) and chicken (NP_001239072.1.) sequences.
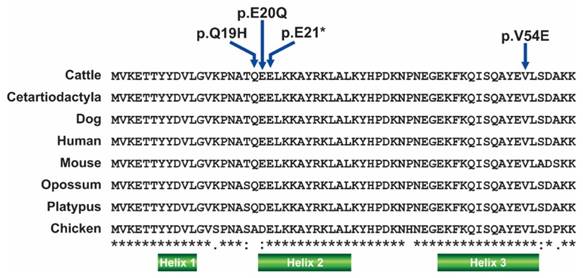
Identified SNPs includes 3 non-synonymous ones (c. 57A>C; p. Q19H, c.58 G>C, p. E20Q; c.161 T>A, p. V54Q), a nonsense mutation (c. 61 G>T, p. E21X) and a silent mutation (c.162 G>A, p. V54). In the American breed, only missense mutation (c.58 G>C, p. E20Q) and silent mutation (c.162 G>A, p. V54) occurred in exon 2 and exon 3 respectively. Alignment of the amplified N terminal coding region with 1hdj (NMR solution structure of the human hsp40 (hdj-1) J domain) suggested that these p. Q19H, p. E20Q and p. E21X amino acid changes occurred in helix 2 while p. V54Q occurred in helix 3 of the J domain. The 3 missense mutations observed in the N terminal, J domain include: (1) c. 57A>C, resulting in an amino acid change from glutamine to histidine; (2) c.58 G>C, resulting in an amino acid change from glutamic acid to glutamine; (3) c.161 T>A resulting in an amino acid change from valine to glutamine. At position 19, glutamine was observed to be conserved in human, cattle, dog, and mouse but was replaced by arginine in chickens. Although amino acid positions 20, 21 and 54 appears to be conserved in cattle, dog, human, mouse and chicken (Figure 3), mutations at these positions observed in this study could plausibly have functional consequences.
In-silico functional analysis using PANTHER predicted a moderately high subPSEC (-2.71504) and Pdeleterious (0.42924) values for c. 57A>C non-synonymous mutation and a much higher subPSEC (-5.83501) and Pdeleterious (0.94454) values for c. 161T>A non-synonymous mutation. Although amino acid changes p. Q19H occurred in helix 2 and p. V54Q in helix 3, putative functional impacts on the protein were much more pronounced in p. V54Q. These putative deleterious amino acid changes (p. Q19H in helix 2 and p. V54Q in helix 3) were only observed in African and Asian cattle breeds but absent in American breeds. As for the amino acid changes observed in yak with respect to the C-terminal region, the amino acid altering mutations were predicted by PANTHER to be non- deleterious, given the Pdeleterious and subPSEC values (0.44, -2.77 and 0.414, -2.65) for p. I310F and p. Y311H amino acid changes respectively [25].
Eleven, 9 and 2 haplotypes identified in Asian (from Pakistan), African (from Nigeria) and American cattle breeds are shown in Tables 1, 2, 3 along with haplotype frequencies. Among Asian breeds, 43.1% of the breeds belonged to haplotype 1 (AGGTG) while the remaining individuals belonged to other haplotypes (Table 1). Surprisingly, while most of the Asian breeds belong to haplotype 1 including yak, Bhagnari breed, a draft type breed with compact and well-proportioned body and limbs lacked haplotype 1. Among African breeds, 40% of individuals belonged to haplotype 1 (AGGTG) (Table 2) while others belonged to other haplotypes. Approximately 85% of American breed (Brangus) belonged to haplotype 1 (Table 3), suggesting this haplotype may be the ancestral haplotype at this locus, for all breeds.
Haplotype frequencies at the DNAJA1 locus in Asian (Pakistani) cattle
| Haplotypea | Achai | Cholistani | Dajal | Dhanni | Lohani | Narimaster | Red Sindhi | Sahiwal | Therparker | Yak | Bhagnari | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AGGTG | 2(0.40) | 4(0.80) | 6(0.86) | 4(0.57) | 1(0.20) | 1(0.25) | 1(0.20) | 2(0.40) | 3(0.50) | 1(0.25) | * | 25 |
| AGGAA | 1(0.20) | 1(0.20) | * | * | * | 1(0.25) | * | * | * | * | * | 3 |
| ACGAG | 1(0.20) | * | * | 1(0.143) | * | * | * | * | * | * | * | 2 |
| AGGTA | 1(0.20) | * | * | 1(0.143) | * | * | * | * | * | * | * | 2 |
| AGTAG | * | * | * | * | * | * | * | * | * | * | 1(0.17) | 1 |
| CGTTG | * | * | * | * | * | * | * | * | * | * | 1(0.17) | 1 |
| AGGAG | * | * | * | 1(0.143) | 2(0.40) | 1(0.25) | * | 1(0.20) | * | 2(0.50) | 2(0.33) | 9 |
| CGGAA | * | * | * | * | * | * | * | 1(0.20) | * | * | 1(0.17) | 2 |
| ACGAA | * | * | 1(0.14) | * | * | * | 2(0.40) | 1(0.20) | 1(0.17) | * | 1(0.17) | 6 |
| ACTTG | * | * | * | * | 1(0.2) | * | 2(0.40) | * | 1(0.17) | 1(0.25) | * | 5 |
| ACTTG | * | * | * | * | 1(0.2) | * | * | * | 1(0.17) | * | * | 2 |
| 58 |
a Haplotypes are indicated following the SNP position in the DNAJA1 gene: c. 57A>C; c. 58G>C; c. 61 G>T; c. 161 T>A; c. 162 G>A. * represents individuals that do not have a particular haplotype
Haplotype frequencies at the DNAJA1 locus in African (Nigerian) cattle
| Haplotypea | Muturu | N'dama | Sokoto Gudali | White Fulani | Total |
|---|---|---|---|---|---|
| AGGTG | 3(0.60) | 2(0.33) | 1(0.20) | 2(0.50) | 8 |
| ACTTG | 1(0.20) | * | * | 1(0.25) | 2 |
| ACGAA | 1(0.20) | * | 2(0.40) | 1(0.25) | 4 |
| ACGTG | * | 1(0.16) | * | * | 1 |
| AGGTA | * | 1(0.16) | * | * | 1 |
| CCTTG | * | 1(0.16) | * | * | 1 |
| AGGAG | * | 1(0.16) | * | * | 1 |
| CGGTG | * | * | 1(0.20) | * | 1 |
| ACTAG | * | * | 1(0.20) | * | 1 |
a Haplotypes are indicated following the SNP position in the DNAJA1 gene: c. 57A>C; c. 58G>C; c. 61 G>T; c. 161 T>A; c. 162 G>A. * represents individuals that do not have a particular haplotype
Haplotype frequencies at the DNAJA1 locus in American cattle
| Haplotypea | Brangus |
|---|---|
| GG | 6(0.857) |
| CA | 1(0.143) |
a Haplotypes are indicated following the SNP position in the DNAJA1 gene: c. 58G>C and c. 162 G>A
Discussion
Hsp40 is known to possess functional domains that play crucial roles in chaperoning non-native proteins, and consists of a conserved J domain that enables Hsp70 to perform its essential cellular functions [26,27]. The expression of DNAJA1, a homolog of Escherichia coli DnaJ, has been implicated in meat tenderness [28]. Our BlastN analysis of the amplified N-terminal J domain and the C-terminal regions of the DNAJA1 gene showed 99% and 100% similarity (except for yak C- terminal region) respectively to the bovine DNAJA1 gene with accession number NM_001015637.1. Five polymorphic loci located in exons 2 and 3 were identified in the N-terminal region (containing the J domain), while intron 2 remained conserved among cattle breeds. Conservation of intron 2 observed in our study, as published [29], may be indicative of its transcriptional role in DNAJA1 gene expression and splicing.
Five polymorphic loci were observed in the N-terminal J domain with the first three mutations (c. 57A>C, c.58 G>C, c. 61 G>T) occurring within 5 bp region in exon 2 while the remaining two mutations (c.161 T>A, c.162 G>A) occurred within 2 bp in exon 3, and these were only observed in Asian and African breeds. Interestingly, the number of mutations observed in American Brangus raised in temperate environment is somewhat different from cattle raised in tropical environments. Although the observed genetic differences might be concomitant with breed population history, whether the mutations are environmentally-induced to plausibly improve the functional activity of DNAJA1 gene in these cattle breeds remain obscure and are subject to further investigation.
The conserved nature of the amplified C-terminal region among domestic cattle and yak irrespective of their geographical differences might suggest a critical role in binding non-native client proteins to avoid deleterious effects of mutations in this region [27]. C-terminal region in DnaJ proteins has been reported to be functionally involved in binding of client proteins [10] and in dimerization and chaperone activity [30]. Also, the C-terminal region possess two-barrel topology domains: the C-terminal domain I (CTD I) and CTD II, in which CTD I has a hydrophobic pocket that is thought to bind client proteins, while the extreme C terminus (CTD II) increases affinity for client proteins through a dimerization domain [31]. The alignment of the consensus sequence of the carboxylic region in domestic cattle and yak showed some subtle variation within DNAJA1 gene and the exonic mutations included c. 914A>G in exon 7, c. 1035A>T and c.1038T>C in exon 8 as well as mutations in the introns. Deletions and substitutions observed in intron 7 and a 124 bp insertion in intron 8 of the C-terminal region may reflect considerable evolutionary divergence between domestic cattle and yak dating up to 5 Mya [32,33]. Whether the 124 bp insertion observed in yak DNAJA1 contributes to the stability of DNAJA1 gene in the yak genome is unknown now, and the functional or structural relevance of this insertion deserves further investigation.
Eleven and 9 haplotypes were identified in Asian and African breeds of cattle with American Brangus (indicine Brahman x taurine Angus) population having two haplotypes. The disparity between the tropical breeds and temperate breeds of cattle with respect to the number of haplotypes could be explained in part by sample size. While it was surprising that c.58 G>C in exon 2 and c.162 G>A in exon 3 were only observed in American Brangus compared to the five polymorphic loci observed in this study in tropical Asian and African breeds, it may support the hypothesis that these genetic differences may have been sculpted by evolutionary forces to improve functional capability in both tropical and temperate environments.
DNAJA1 possesses the same structural features that are found in E. coli DnaJ [13]. These features include a J domain, a 70-amino acid sequence of 4 helices and a loop region linking helices II and III with a HPD motif [16], a glycine-phenylalanine (G/F)-rich region, 2 zinc-finger-like motifs, and a conserved carboxyl-terminal domain (CTD). In this study, mutations were observed in positions 19, 20 and 21, which were earlier predicted to be part of the amino acids found in helix 2. In-silico functional analysis of these SNP positions suggested no deleterious effects of these mutations based on their Pdeleterious and their subPSEC values. The reasons for the non-deleterious effects of these mutations could be due to the important functional roles played by helix 2 in Hsp70 ATPase activity.
Genetic studies have proposed that several DnaK mutations exists, thereby suppressing the effects of mutations in the critical HPD motif of DnaJ [18]. Whether effects of mutations in the helix region of the J domain are suppressed by mutations in DnaK is unclear, but a published report [34] observed that substitution of certain key residues in helix 2 disrupts J domain-based interactions with Hsp70. In-silico functional analysis of the missense p. V54Q mutation predicted much higher deleterious effects with a likely functional consequence. To this end, we recommend additional experiments involving site-directed mutagenic studies to evaluate the effects of these mutations on their chaperoning functions, to verify present speculations.
Differences in the C-terminal region SNPs between yak and domestic cattle may be due to speciation effects. The missense mutations (p. I310F and p. Y311H) observed in yak was predicted by PANTHER to be non-deleterious, further supporting our previous assertion that evolutionary avoidance of mutations in this region is imperative for the functional role of binding to client proteins. The results from our study underscore the need for more work in this area since our analysis focused on the sequences of the N-terminal and C-terminal regions while taking cognizance of the important roles played by other regions (G/F and the zinc-finger domains) in chaperoning functions. Complete genomic analysis of the entire DNAJA1 sequence and the analysis of functional effects of DNAJA1 polymorphisms on prevention of protein misfolding, heat tolerance and meat tenderness are areas ripe for future investigations. These will enable us gain better understanding of the functional consequences of DNAJA1 gene variants on economically important traits in cattle and other livestock species.
In conclusion, differences in DNAJA1 (Hsp40) gene exists between tropical and temperate cattle from Africa, Asia and America, but the number of mutations observed in American Brangus raised in temperate environment was different from cattle raised in tropical environments. These differences may have been sculpted by evolutionary forces during adaptation of these cattle breeds. The conservation of the carboxylic terminal region in domestic cattle and yak suggest its critical role in binding non-native proteins. However, the 124 bp insertion observed in yak may have functional or structural roles that are yet unknown. Taken together, our results support significant diversity at this locus that may have functional impact on climate adaptation in animals.
Acknowledgements
We are grateful for the financial support from the College of Agriculture and Life Sciences, Cornell University, Ithaca, NY and Zoetis, Inc. Additional support by National Research Initiative Competitive Grant Program (Grant No. 2006-35205-16864) from the USDA National Institute of Food and Agriculture, USDA-NIFA Research Agreements (Nos. 2009-65205-05635, 2010-34444-20729) and USDA Federal formula Hatch funds appropriated to the Cornell University Agricultural Experiment Station are gratefully acknowledged. OOA was supported by a 1278468 fellowship from the US Agency for International Development; BNT is supported by a Laboratory Support and Faculty Development Award, College of Health Sciences and Technology, Rochester Institute of Technology. Visiting fellowships awarded to TH and WAK by the Higher Education Commission of Pakistan are also gratefully acknowledged.
Author contributions
OOA, MDD, IGI and BNT conceived the project; OOA, SOP, MDD and IGI designed and performed the experiments; OOA, SOP, MDD, IGI, BNT analyzed the data; OOA, IGI, BNT wrote the manuscript; FDM, WAK, TH, MEB contributed to the scientific content. All authors read and approved the final manuscript.
Competing Interests
The authors have declared that no competing interest exists.
References
1. Hartl FU, Hayer-Hartl M. Molecular chaperones in the cytosol, from nascent chain to folded protein. Science. 2000;295:1852-1858
2. Harris JM, Nguyen PP, Patel MJ, Sporn ZA, Hines JK. Functional diversification of hsp40: distinct j-protein functional requirements for two prions allow for chaperone-dependent prion selection. PLoS Genet. 2014;10(7):e1004510
3. Kim JH, Alderson TR, Frederick RO, Markley JL. Nucleotide-dependent interactions within a specialized Hsp70/Hsp40 complex involved in Fe-S cluster biogenesis. J Am Chem Soc. 2014;136(33):11586-9
4. Rodriguez KA, Osmulski PA, Pierce A, Weintraub ST, Gaczynska M, Buffenstein R. A cytosolic protein factor from the naked mole-rat activates proteasomes of other species and protects these from inhibition. Biochim Biophys Acta. 2014;1842(11):2060-72
5. Koutras C, Braun JE. 2014. J protein mutations and resulting proteostasis collapse. Front Cell Neurosci. 2014;8:191
6. Stein KC, Bengoechea R, Harms MB, Weihl CC, True HL. Myopathy-causing mutations in an HSP40 chaperone disrupt processing of specific client conformers. J Biol Chem. 2014;289(30):21120-21130
7. Hennessy F, Nicoll W.S, Zimmermann R, Cheetham M.E. and Blatch G.L. Not all J domains are created equal: Implications for the specificity of hsp40-hsp70 interaction. Protein Sci. 2005;14:1697-1709
8. Baaklini I, Wong MJ, Hantouche C, Patel Y, Shrier A, Young JC. The DNAJA2 substrate release mechanism is essential for chaperone-mediated folding. J Biol Chem. 2012;287(50):41939-54
9. Lee S, Fan CY, Younger JM, Ren HY, Cyr DM. Identification of essential residues in the Type II Hsp40 Sis1 that function in polypeptide binding. J Biol Chem. 2002;277:21675-21682
10. Li JZ, Oian XG, Sha B. The crystal structure of the yeast Hsp40 Ydj1 complexed with its peptide substrate. Structure. 2003;11:1475-1483
11. Langer T, Lu C, Echols H, Flanagan J, Hayer MK. et al. Successive action of Dnak, Dnaj and Groel along the pathway of chaperone-mediated protein folding. Nature. 1992;356:683-689
12. Li J, Qian XG, Sha BD. Heat shock protein 40: structural studies and their functional implications. Protein Pept Lett. 2009;16:606-612
13. Cheetham ME. and Caplan AJ. Structure, function and evolution of DnaJ: Conservation and adaptation of chaperone function. Cell Stress Chaperones. 1998;3:28-36
14. Szyperski T, Pellecchia M, Wall D, Georgopoulos C, Wuthrich K. NMR Structure determination of the Escherichia coli Dnaj molecular chaperone - secondary structure and backbone fold of the N-terminal region (residues-2-108) containing the highly-conserved J-domain. Proc Natl Acad Sci USA. 1994;91:11343-11347
15. Wittung-Stafshede P, Guidry J, Horne BE, Landry SJ. The J domain of Hsp40 couples ATP hydrolysis to substrate capture in Hsp70. Biochemistry. 2003;42:4937-4944
16. Qian YQ, Patel D, Hartl FU, McColl DJ. Nucleic Magnetic Resonance solution structure of the human Hsp40 HDJ-1 J domain. J Mol Biol. 1996;260:224-235
17. Greene M, Makos K, Landry SJ. Role of the J-domain in the cooperation of Hsp40 with Hsp70. Proc Natl Acad Sci USA. 1998;95:6108-6113
18. Berjanskii MV, Riley MI, Van Doren SR. Hsc70-interacting HPD loop of the J domain of polyomavirus T antigens fluctuates in ps to ns and µs to ms. J Mol Biol. 2002;321:503-516
19. Landry SJ. Structure and energetics of an allele-specific interaction between dnaJ and dnaK: Correlation of nuclear magnetic resonance chemical shift perturbations in the J-domain of Hsp40/DnaJ with binding affinity for the ATPase domain of Hsp70/DnaK. Biochemistry. 2003;42:4926-4936
20. Sha BD, Lee S, Cyr DM. The crystal structure of the peptide-binding fragment from the yeast Hsp40 protein Sis1. Structure. 2000;8:799-807
21. Adefenwa MA, Peters SO, Agaviezor BO, Wheto M, Adekoya KO, Okpeku M, Oboh B, Williams GO, Adebambo OA, Singh M, Thomas B, De Donato M, Imumorin IG. Identification of single nucleotide polymorphisms in the agouti signaling protein (ASIP) gene in some goat breeds in tropical and temperate climates. Mol Biol Rep. 2013;40:4447-57
22. Rozas J, Sánchez-DelBarrio JC, Messeguer X, Rozas R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics. 2003;19(18):2496-2497
23. Thomas PD, Campbell MJ, Kejariwal A. et al. PANTHER: A library of protein families and subfamilies indexed by function. Genome Res. 2003;13:2129-2141
24. Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. ClustalW and ClustalX version 2. Bioinformatics. 2007;23(21):2947-2948
25. Brunham LR, Singaraja RR, Pape TD, Kejariwal A, Thomas PD, Hayden MR. Accurate prediction of the functional significance of single nucleotide polymorphisms and mutations in the ABCAI gene. PLoS Genet. 2005;1:e83
26. Sahi C. and Craig EA. Network of general and specialty J protein chaperones of the yeast cytosol. Proc Natl Acad Sci USA. 2007;104:7163-7168
27. Sarraf NS, Shi R, McDonald L, Baardsnes J, Zhang L, Cygler M, Ekiel I. Structure of CbpA J-domain bound to the regulatory protein CbpM explains its specificity and suggests evolutionary link between CbpM and transcriptional regulators. PLoS One. 2014;9(6):e100441
28. Bernard C, Cassar-Malek I, Le Cunff M, Dubroeucq H, Renand G, Hocquette JF. New indicators of beef sensory quality revealed by expression of specific genes. J Agric Food Chem. 2007;55:5229-5237
29. Marty A, Amigues Y, Servin B, Renand G, Leveziel H, Rocha D. Genetic variability and linkage disequilibrium patterns in the bovine DNAJA1 gene. Mol Biotechnol. 2010;44:190-197
30. Kota P, Summers DW, Ren HY, Cyr DM, Dokholyan NV. Identification of a consensus motif in substrates bound by a Type I Hsp40. Proc Natl Acad Sci USA. 2009;106:11073-11078
31. Wu Y, Li J, Jin Z, Fu Z, Sha B. The crystal structure of the C-terminal fragment of yeast Hsp40 Ydj1 reveals novel dimerization motif for Hsp40. J Mol Biol. 2005;346:1005-1011
32. Bradley DG, MacHugh DE, Cunningham P, Loftus RT. Mitochondrial diversity and the origins of African and European cattle. Proc Natl Acad Sci USA. 1996;93(10):5131-35
33. Ritz LR, Glowatzki-Mullis ML, MacHugh DE, Gaillard C. Phylogenetic analysis of the tribe Bovini using microsatellites. Anim Genet. 2000;31(3):178-85
34. Suh WC, Burkholder WF, Lu CZ, Zhao X, Gottesman ME, Gross CA. Interaction of the Hsp70 molecular chaperone, DnaK, with its cochaperone DnaJ. Proc Natl Acad Sci USA. 1998;95:15223-15228
Author contact
![]() Corresponding authors: Dr. Bolaji N. Thomas, Department of Biomedical Sciences, Rochester Institute of Technology, Rochester NY, 14623. USA; Tel: +1 585-475-6382; E-mail: bntsbiedu. Dr. Ikhide G. Imumorin, School of Biological Sciences, Georgia Institute of Technology, Atlanta, GA 30332. USA; Tel: +1 404-894-3747; E-mail: igi2gatech.edu.
Corresponding authors: Dr. Bolaji N. Thomas, Department of Biomedical Sciences, Rochester Institute of Technology, Rochester NY, 14623. USA; Tel: +1 585-475-6382; E-mail: bntsbiedu. Dr. Ikhide G. Imumorin, School of Biological Sciences, Georgia Institute of Technology, Atlanta, GA 30332. USA; Tel: +1 404-894-3747; E-mail: igi2gatech.edu.